The 5 Assumptions of Hardy-Weinberg Equilibrium

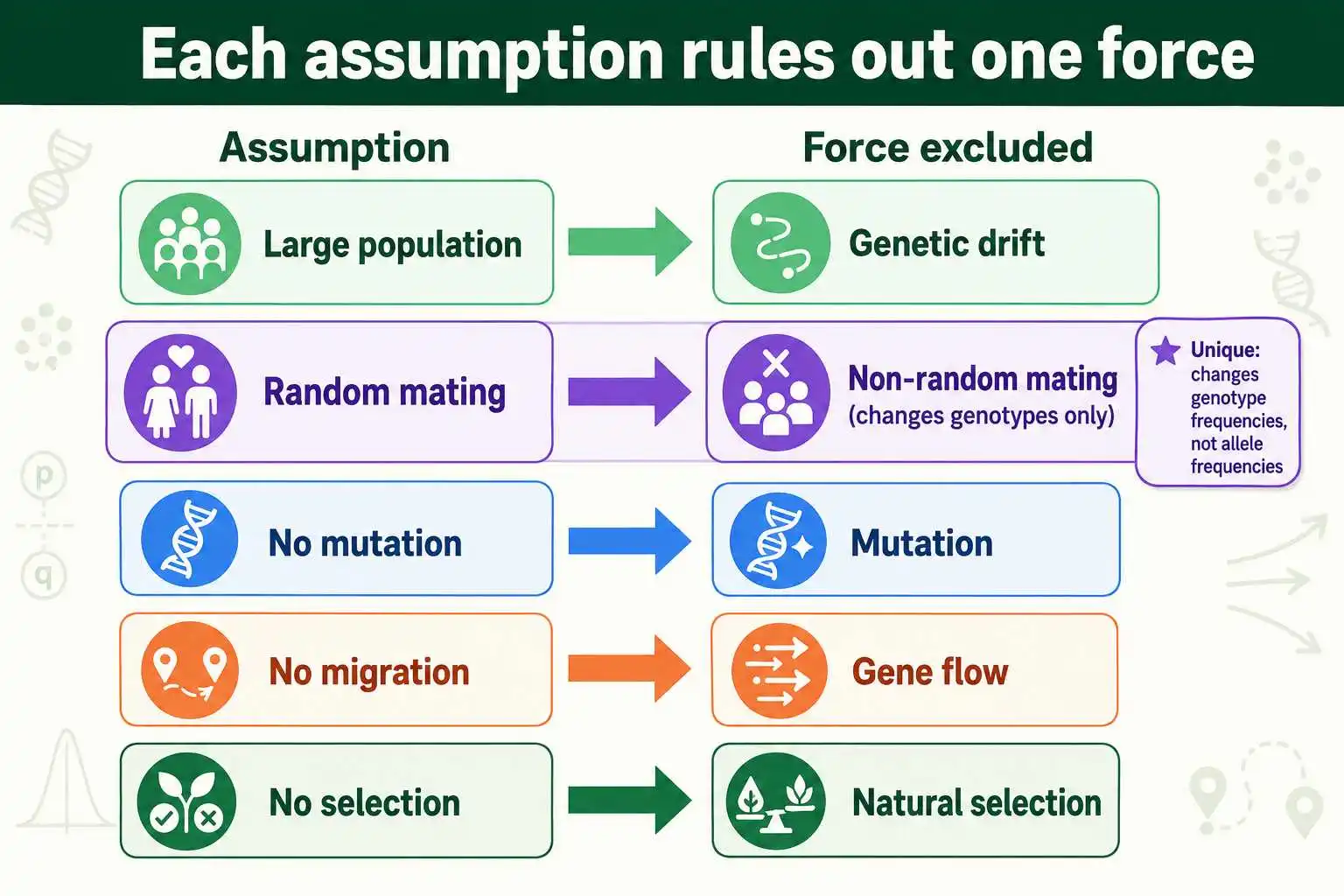
Hardy-Weinberg equilibrium holds only when a population meets five assumptions: a large population size, random mating, no mutation, no migration, and no natural selection. Each one rules out a specific force that would otherwise change allele or genotype frequencies. When all five hold, the population's genetic makeup stays constant across generations. When any one is violated, the population departs from equilibrium, which is the signal that evolution is occurring at that gene.
These five assumptions are the conditions baked into the Hardy-Weinberg model, and understanding each one is essential for using the principle correctly. This guide explains all five, what each prevents, and exactly what happens when each is violated. It also covers a subtle but important distinction: one of the five affects genotype frequencies without touching allele frequencies, which sets it apart from the others. The Hardy-Weinberg equation can be applied with a calculator, but knowing the assumptions tells you when the prediction can be trusted.
Why the Assumptions Matter
Before going through them one by one, it helps to understand why these assumptions exist at all. The Hardy-Weinberg principle is a mathematical model, and like any model, it makes simplifying assumptions to produce a clean result. The five assumptions describe an idealized population in which nothing disturbs the genetic frequencies.
The deeper meaning is that each assumption corresponds to one of the forces that cause evolution. Evolution, at the genetic level, is a change in allele frequencies over time. The Hardy-Weinberg model strips away every force that could cause such a change, leaving a population frozen in genetic stillness. So the five assumptions are really a list of the five things that must be absent for a population not to evolve. Turn that around, and the same list names the five forces that drive evolutionary change.
This is why the assumptions are so useful despite describing a situation that never perfectly exists. No real population meets all five conditions exactly, which means the equilibrium is a theoretical baseline rather than a description of reality. Its value lies in comparison: by checking whether a population matches the equilibrium prediction, scientists can detect when one of the assumptions is being violated and therefore when evolution is happening. The assumptions, in other words, are the bridge between the static model and the dynamic reality of evolving populations. The full logic of this null-model role is explained in our guide on what Hardy-Weinberg equilibrium is.
Assumption 1: A Large Population
The first assumption is that the population must be very large, ideally infinite in the theoretical model. This condition exists to rule out genetic drift, the random change in allele frequencies that happens by chance in finite populations.
The logic comes from probability. In a large population, random variation in which individuals reproduce averages out, so allele frequencies stay stable from generation to generation. In a small population, chance plays a much bigger role. If only a few individuals reproduce, the alleles they happen to pass on may not reflect the population's true frequencies, and the frequencies can drift purely by luck. Imagine flipping a coin ten times versus ten thousand times: the small sample can easily stray far from fifty-fifty, while the large sample reliably approaches it. Allele frequencies behave the same way.
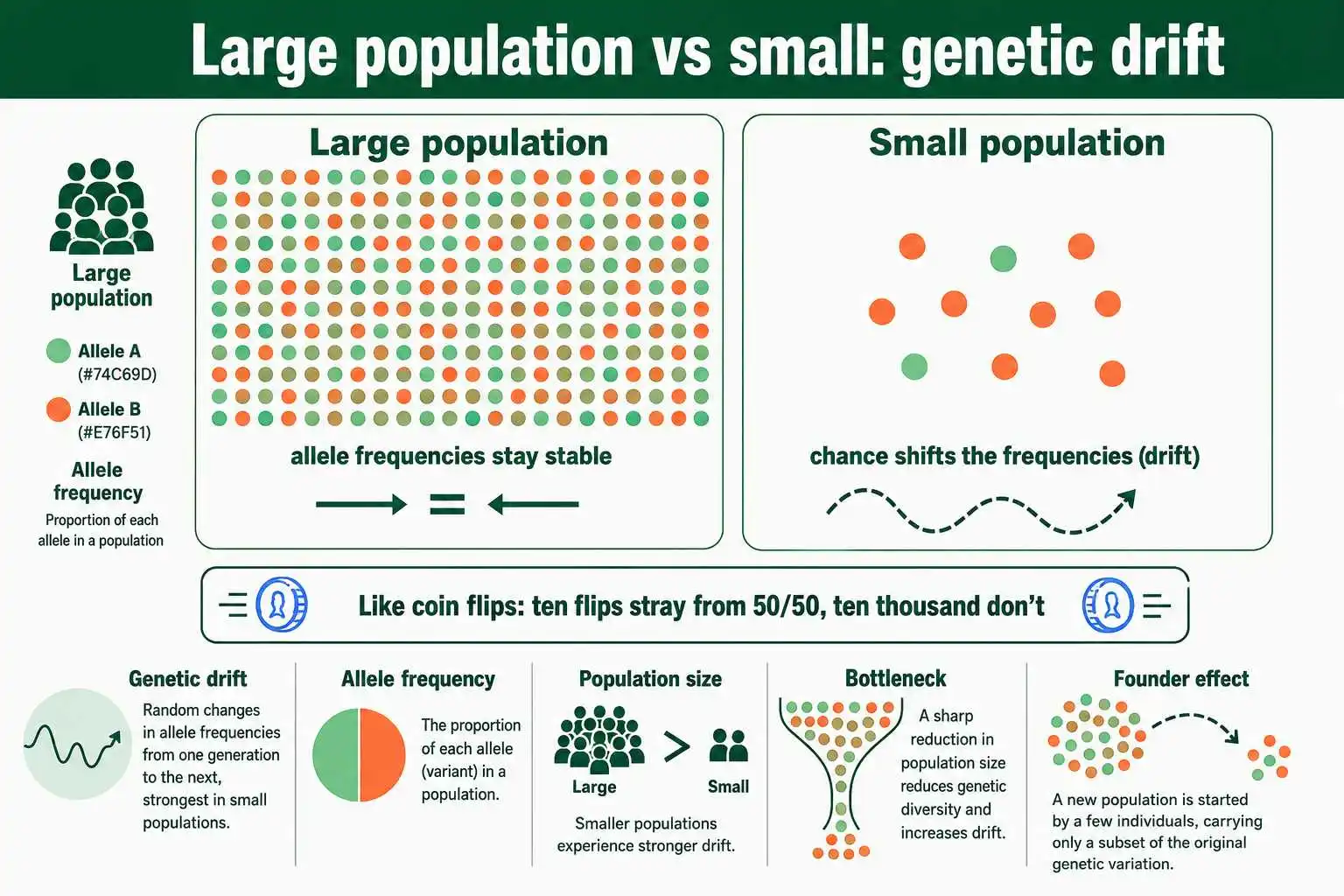
When this assumption is violated, in a small population, genetic drift causes allele frequencies to fluctuate randomly, and alleles can even be lost entirely or become fixed by chance alone. This is a real evolutionary force, especially important in small or isolated populations, and it operates without any regard to whether an allele is beneficial or harmful. A large population size is therefore the condition that keeps random chance from reshaping the gene pool. Because no real population is infinite, every actual population experiences some drift, though the effect is negligible in very large populations and dominant in very small ones.
Two well-known situations make this concrete. A population bottleneck occurs when a population's size crashes dramatically, perhaps from disease or habitat loss, so that the survivors carry only a fraction of the original genetic variation, and their allele frequencies may differ sharply from the original population purely by chance. The cheetah is a classic example, having passed through a severe bottleneck that left the species with strikingly low genetic diversity. A related case is the founder effect, where a small group splits off to start a new population, carrying only a non-representative sample of the parent population's alleles. Both show how a small population size lets chance, rather than fitness, dictate which alleles persist, exactly the situation the large-population assumption is designed to exclude.
Assumption 2: Random Mating
The second assumption is random mating, meaning individuals pair without regard to their genotype at the gene in question. Every individual has an equal chance of mating with any other, so allele combinations in the offspring form purely by chance.
This assumption is special, and understanding why reveals something important about the model. Random mating is the condition that produces the p², 2pq, q² genotype ratios in the first place. The squared terms come from combining alleles at random, exactly as you would multiply probabilities, so without random mating, the genotype frequencies need not match the Hardy-Weinberg prediction even if the allele frequencies are unchanged. The genotype distribution depends directly on how mating occurs.
When random mating is violated, the effect is distinctive and sets it apart from the other four assumptions. Non-random mating takes several forms. Inbreeding is mating between relatives, who are more likely to share the same alleles, which raises the frequency of homozygotes. Assortative mating is when individuals choose partners similar to themselves for a trait, such as size or coloration, which similarly concentrates matching alleles together. The opposite, disassortative mating, where unlike individuals pair, increases heterozygotes instead. In each case the pattern of pairing, not chance, determines which genotypes form. Non-random mating changes genotype frequencies without changing allele frequencies. This is the crucial subtlety: forms of non-random mating like inbreeding rearrange how alleles are packaged into genotypes, typically increasing homozygotes and decreasing heterozygotes, but the overall proportion of each allele in the gene pool stays the same. The alleles are the same; they are just combined differently. This makes non-random mating unique among the five, because the other four assumptions, when violated, actually change allele frequencies, while non-random mating alone changes only the genotype frequencies.

Assumption 3: No Mutation
The third assumption is that there is no mutation. Mutation is the process that creates new alleles by changing the DNA sequence, and the model assumes this is not happening, so the set of alleles and their frequencies are not altered by new genetic changes.
Mutation matters to equilibrium because it is the ultimate source of all genetic variation. Every allele that exists originated as a mutation at some point. If mutations occur during the timeframe being considered, they introduce new alleles or convert one allele into another, directly changing allele frequencies. The Hardy-Weinberg model assumes no such changes occur, so the alleles present at the start remain the only alleles, in their original proportions.
When this assumption is violated, mutation gradually changes allele frequencies, though usually very slowly. Mutation rates are typically low, so the effect on allele frequencies over a single generation is tiny, far smaller than the effects of selection or drift in most cases. This is why mutation is often considered a weak force for changing frequencies in the short term, even though it is supremely important over long timescales as the origin of all variation. For the purposes of a Hardy-Weinberg calculation spanning a generation or two, the no-mutation assumption is usually a reasonable approximation, which is part of why the model works as well as it does despite mutation never truly stopping.
There is a useful way to think about mutation's dual role. On the timescale of a few generations, which is what most Hardy-Weinberg problems consider, mutation changes allele frequencies so slowly that ignoring it introduces almost no error. But on the timescale of thousands or millions of generations, mutation is everything, because it is the only process that creates the genetic variation that the other forces then act upon. Selection can favor an allele, but mutation had to produce that allele first. So the no-mutation assumption is safe for short-term prediction yet would be deeply wrong as a statement about evolution over the long run. Holding both ideas at once, mutation as negligible in the short term but foundational over deep time, captures its true place in population genetics.
Assumption 4: No Migration
The fourth assumption is no migration, also called no gene flow. This means no individuals enter or leave the population, so no alleles are added or removed by movement between populations.
Migration affects allele frequencies because incoming or outgoing individuals carry alleles with them. If individuals from a population with different allele frequencies migrate in, they bring their alleles, shifting the frequencies of the receiving population toward those of the source. Likewise, individuals leaving take their alleles with them. This flow of alleles between populations, called gene flow, tends to make separate populations more genetically similar over time, because it mixes their gene pools.
When this assumption is violated, gene flow changes allele frequencies in both the source and receiving populations. It is an important evolutionary force, especially for populations that are not fully isolated, which is most real populations. Gene flow can introduce entirely new alleles to a population or alter the balance of existing ones, and it counteracts the genetic divergence that drift and local selection would otherwise produce. The no-migration assumption requires a closed population with no genetic exchange, which is why truly isolated populations, like those on remote islands, come closer to meeting it than populations connected to their neighbors.
Assumption 5: No Natural Selection
The fifth assumption is no natural selection, meaning all genotypes have equal rates of survival and reproduction. No genotype confers any advantage or disadvantage, so no allele is favored or disfavored, and the frequencies stay constant.
Natural selection is probably the most famous evolutionary force, and it directly changes allele frequencies whenever genotypes differ in fitness. If individuals with one genotype survive better or reproduce more, they pass on more copies of their alleles, so those alleles become more common over generations. The Hardy-Weinberg model assumes this does not happen, that every genotype is equally successful, which keeps allele frequencies fixed. This assumption also covers sexual selection, where mate choice based on traits would favor certain alleles.
When this assumption is violated, selection systematically changes allele frequencies in the direction of higher fitness, which is the core mechanism of adaptive evolution. Unlike drift, which is random, selection is directional, consistently favoring whichever alleles confer an advantage in the current environment. This makes it one of the most powerful forces shaping real populations. There is a subtle case worth noting: some forms of selection, like balancing selection that maintains multiple alleles, can actually keep a population looking close to Hardy-Weinberg equilibrium even though selection is occurring, which is why a population matching the prediction does not absolutely guarantee that no selection is happening. The detailed workings of all these forces are covered in our guide on what disrupts Hardy-Weinberg equilibrium.
The Often-Forgotten Technical Assumptions
Beyond the five main assumptions, the Hardy-Weinberg model quietly rests on a few additional technical conditions that textbooks often gloss over. They are worth knowing, because they define the kind of organism and gene the model applies to in the first place.
The first is that the organism is diploid, carrying two copies of each gene, one from each parent. The entire p² + 2pq + q² framework depends on individuals having exactly two alleles per gene, which is what makes the binomial expansion of (p + q)² describe the genotypes. The model does not directly apply to haploid organisms or to genes with different ploidy without modification. The second technical assumption is that reproduction is sexual, since the model describes how alleles recombine when gametes from two parents unite. Asexual reproduction would not shuffle alleles in the way the equation assumes.
A third technical assumption is that generations do not overlap, meaning the model treats reproduction as happening in discrete cycles rather than continuously. This keeps the math clean, letting one generation be compared neatly to the next. A fourth, sometimes stated separately, is that the gene under study is autosomal rather than sex-linked, because sex-linked genes follow a different distribution between males and females and need a modified treatment. These technical assumptions are usually satisfied automatically for the textbook cases the principle is applied to, which is why they receive less attention than the five main conditions. But they are part of the model's foundation, and recognizing them clarifies exactly what kind of situation Hardy-Weinberg describes.
How a Violated Assumption Is Detected
Knowing the assumptions is one thing; recognizing when one has been violated in real data is another, and it is where the principle becomes a practical tool. Each violated assumption tends to leave a characteristic fingerprint in the genotype frequencies.
The general method is to compare observed genotype frequencies against the frequencies the Hardy-Weinberg equation predicts from the allele frequencies. If they match closely, the population may be in equilibrium for that gene. If they differ significantly, an assumption is being violated and the deviation often hints at which one. A clear example is an excess of homozygotes and a shortage of heterozygotes compared to the prediction, which is the classic signature of inbreeding or population structure, both forms of non-random mating. The allele frequencies can be unchanged while the genotype distribution is visibly skewed toward homozygotes.
Other violations leave different marks. A steady directional change in an allele's frequency across generations points to selection or gene flow rather than random mating effects. Random, undirected swings in a small population suggest genetic drift. The formal way to test whether observed and expected genotype frequencies differ more than chance allows is a chi-square test, which converts the comparison into a statistical decision, and we devote a full guide to testing for Hardy-Weinberg equilibrium with chi-square. One important caution applies, though: matching the prediction does not absolutely prove no force is acting, because certain situations, like balancing selection, can produce equilibrium-looking frequencies even while selection operates. Deviation reliably indicates a violated assumption, but conformity is suggestive rather than conclusive.
A Memory Aid for the Five Assumptions
Because the five assumptions come up constantly, especially in exams, a mnemonic helps lock them in. A popular one is "Big Moose Never Run South," where each word cues one assumption.
The mnemonic maps cleanly onto the five conditions. Big stands for a big population, ruling out genetic drift. Moose cues no Mutation. Never cues No migration, that is, no gene flow. Run cues Random mating. South cues no Selection. Running through the phrase lets you recall all five conditions quickly and in order, which is exactly what an exam question listing the assumptions requires.
It is worth pairing the mnemonic with the deeper understanding, rather than relying on it alone. Each assumption rules out one evolutionary force: drift, mutation, gene flow, non-random mating effects, and selection. If you remember that the assumptions are simply "no evolutionary forces acting," you can reconstruct the list even without the mnemonic, because you are naming the things that change allele or genotype frequencies. The mnemonic is a useful shortcut for recall, but the concept, that equilibrium means the absence of these forces, is what lets you apply the assumptions correctly to any situation.
Frequently Asked Questions
What are the five assumptions of Hardy-Weinberg equilibrium?
The five assumptions are a large population size, random mating, no mutation, no migration (no gene flow), and no natural selection. Each rules out a force that would change allele or genotype frequencies. When all five hold, the population's genetic frequencies stay constant.
Which assumption affects genotype frequencies but not allele frequencies?
Random mating. When mating is non-random, such as with inbreeding, genotype frequencies change, typically more homozygotes and fewer heterozygotes, but the allele frequencies stay the same. The other four assumptions, when violated, actually change allele frequencies.
Why does Hardy-Weinberg require a large population?
To rule out genetic drift, the random change in allele frequencies that occurs by chance in small populations. In a large population, random variation averages out and frequencies stay stable, while in a small population, chance alone can shift or even fix alleles.
Do any real populations meet all five assumptions?
No real population meets all five perfectly, so true equilibrium is a theoretical ideal. This is by design: the value of the model is as a baseline, and comparing real populations against it reveals which assumptions are violated and therefore which evolutionary forces are at work.
The Five in Perspective
The five assumptions of Hardy-Weinberg equilibrium, large population, random mating, no mutation, no migration, and no selection, together define a population that is not evolving. Each rules out a specific evolutionary force: genetic drift, non-random mating effects, mutation, gene flow, and selection. Four of them change allele frequencies when violated, while random mating uniquely changes only genotype frequencies, leaving the allele frequencies untouched.
Understanding these assumptions is what lets you use Hardy-Weinberg correctly, because they tell you when the model's predictions apply and when a population has departed from equilibrium. The mnemonic "Big Moose Never Run South" helps with recall, but the real insight is that equilibrium is simply the absence of the forces that drive evolution. You can run Hardy-Weinberg predictions with the allele frequency calculator, keeping the assumptions in mind as the conditions under which those predictions hold. For an AP-focused review that uses the "Big Moose Never Run South" mnemonic, this Hardy-Weinberg guide from Albert is a useful reference, and CK-12 offers a concise summary of all five conditions.