Dihybrid Cross: How to Get the 9:3:3:1 Ratio (4×4 Punnett Square)

A dihybrid cross is a genetic cross that follows two traits at once. When you cross two parents that are heterozygous for both traits, the offspring appear in a 9:3:3:1 phenotypic ratio. Nine show both dominant traits, three show the first dominant with the second recessive, three show the reverse, and one shows both recessive traits.
That single ratio, 9:3:3:1, is the headline result of classical genetics with two genes. It looks intimidating next to the simple 3:1 of a single trait, but it comes from the same rules applied twice. This guide shows you how to find the gametes, build the 16-box grid, read the four phenotype classes, and reach the same answer faster with probability. By the end you will be able to solve any two-trait cross by hand and know exactly why the numbers fall where they do.
What Is a Dihybrid Cross?
A dihybrid cross studies the inheritance of two different traits controlled by two different genes. The prefix "di" means two, so you are tracking two characteristics together instead of one. Each gene still has two alleles, a dominant and a recessive form, and each parent still carries two alleles per gene.
The classic example comes from Mendel's pea plants, and it pairs seed shape with seed color. Round (R) is dominant over wrinkled (r). Yellow (Y) is dominant over green (y). A plant that is heterozygous for both traits has the genotype RrYy. Cross two of these plants, RrYy by RrYy, and you get the famous 9:3:3:1 ratio in the next generation.
If a single trait still feels shaky, work through the monohybrid cross and the 3:1 ratio first, because a dihybrid cross is really two monohybrid crosses happening side by side. Everything you know about dominant and recessive alleles, homozygous and heterozygous genotypes, and reading a Punnett square carries straight over. The only thing that changes is the size of the problem.
A dihybrid cross matters because real organisms inherit many traits at once, not one in isolation. Mendel used two-trait crosses to discover his second law, the law of independent assortment, which describes how genes for separate traits behave during reproduction. That law is the reason the ratio comes out to 9:3:3:1 and not something else, and we will get to exactly why later in this guide.
The 9:3:3:1 Ratio at a Glance
Before the mechanics, here is the destination. A dihybrid cross between two double-heterozygotes produces four visible groups of offspring in a fixed ratio.
| Phenotype class | Ratio | Genotype pattern | Pea example |
|---|---|---|---|
| Both dominant | 9 | R_Y_ | Round yellow |
| First dominant, second recessive | 3 | R_yy | Round green |
| First recessive, second dominant | 3 | rrY_ | Wrinkled yellow |
| Both recessive | 1 | rryy | Wrinkled green |
The blank line in a genotype pattern, written R_, means "any second allele," because a single dominant allele is enough to show the dominant trait. So R_Y_ covers every offspring that has at least one R and at least one Y. Nine of the sixteen offspring fit that description, which is why the largest group is the double-dominant one.
Read the ratio as a proportion out of sixteen, since the grid has sixteen boxes. Nine sixteenths show both dominant traits, three sixteenths and another three sixteenths show the two mixed combinations, and one sixteenth shows both recessive traits. Those fractions add up to sixteen sixteenths, the whole population, which is a quick way to check your work.
Setting Up a Dihybrid Cross: Finding the Gametes
Every cross starts with gametes, and this is the step where two-trait problems trip people up. A gamete carries one allele from each gene, never both alleles of either gene. So a RrYy parent does not pass on "RrYy." It passes on a combination that takes one allele from the R gene and one from the Y gene.
Work out every possible combination. From the R gene, a gamete gets either R or r. From the Y gene, it gets either Y or y. Combine those choices every way and you get four gamete types: RY, Ry, rY, and ry. A double-heterozygote always makes four kinds of gamete, each equally likely.
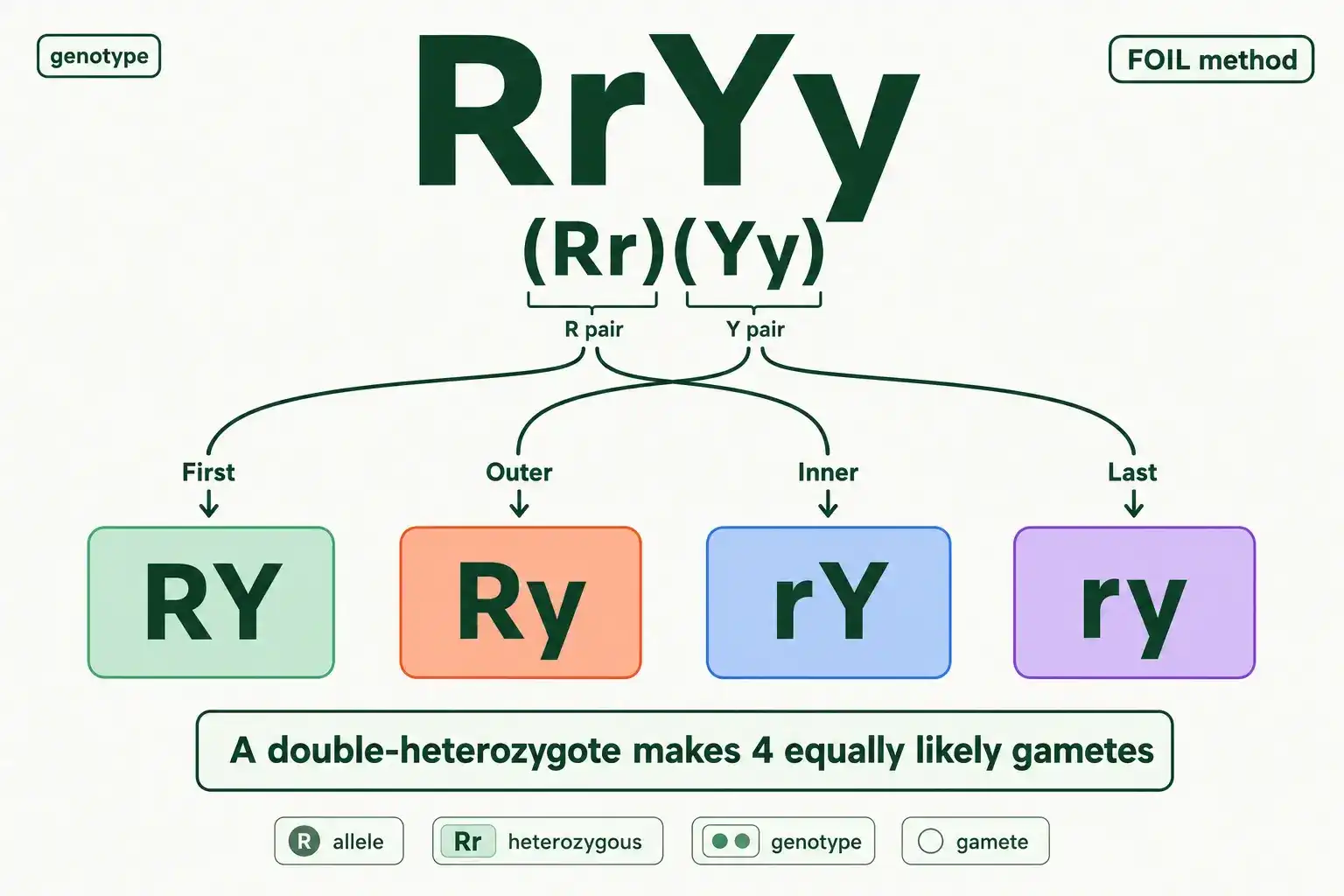
The FOIL method keeps this organized. FOIL stands for first, outer, inner, last, the same trick from multiplying binomials. Treat RrYy as two pairs, (Rr) and (Yy). First pairs the first letters, R and Y, giving RY. Outer pairs R and y, giving Ry. Inner pairs r and Y, giving rY. Last pairs r and y, giving ry. Four gametes, no duplicates, no omissions.
Getting these four gametes right is everything, because they become the labels on both axes of your grid. A single mistake here, like writing "Rr" instead of pulling one allele from each gene, breaks the entire square. Write the four gametes out carefully before you draw anything, and double check that each one has exactly one shape allele and one color allele.
Building the 4×4 Punnett Square
The grid size matches the gamete count. Each parent makes four gametes, so the square is four columns by four rows, giving sixteen boxes in total. Write one parent's four gametes across the top, one per column. Write the other parent's four gametes down the left side, one per row.
Now fill each box by combining its column gamete with its row gamete. A box in the RY column and the rY row, for example, combines RY with rY to give RrYY. Work left to right, top to bottom, and write each genotype with the shape alleles first and the color alleles second so your notation stays consistent.
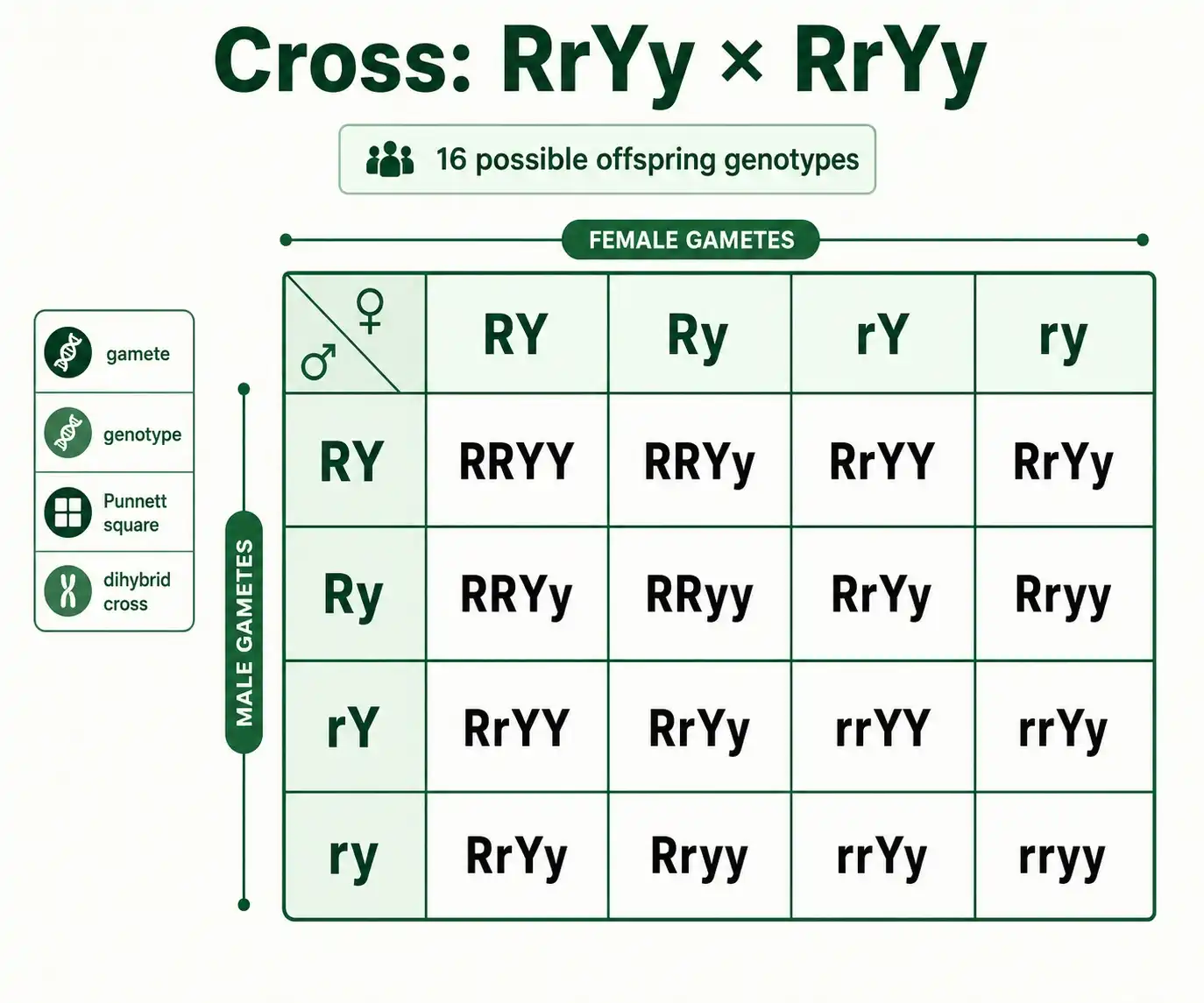
Here is the completed grid for RrYy by RrYy:
| RY | Ry | rY | ry | |
|---|---|---|---|---|
| RY | RRYY | RRYy | RrYY | RrYy |
| Ry | RRYy | RRyy | RrYy | Rryy |
| rY | RrYY | RrYy | rrYY | rrYy |
| ry | RrYy | Rryy | rrYy | rryy |
Sixteen boxes, every possible offspring genotype. Filling them by hand is slow and easy to slip up on, which is exactly the pain a tool removes. The dihybrid cross probability calculator builds this grid instantly and tallies the results, so once you understand the method you can let it handle the bookkeeping.
Reading the 9:3:3:1 Phenotypic Ratio
The grid gives you sixteen genotypes, but you do not have sixteen different appearances. Dominance collapses many genotypes into the same visible trait, which is how sixteen boxes become four phenotype groups.
Sort the boxes by what they show. Any genotype with at least one R and at least one Y looks round and yellow. Count the boxes that fit R_Y_ and you find nine of them. Any genotype with at least one R but two recessive color alleles (R_yy) looks round and green, and there are three. Any genotype that is wrinkled but has at least one Y (rrY_) looks wrinkled and yellow, and there are three. Only the single rryy box is wrinkled and green.

That gives nine round yellow, three round green, three wrinkled yellow, and one wrinkled green, a clean 9:3:3:1 phenotypic ratio. Notice how the two largest single-trait groups overlap in the nine. A plant can be round and yellow through many different genotypes, from the double-dominant RRYY to the double-heterozygote RrYy, and they all land in the same visible class. This is the heart of why phenotype ratios and genotype ratios differ so sharply in a dihybrid cross.
Why 9:3:3:1? The Law of Independent Assortment
The ratio is not a coincidence or a rule to memorize. It follows directly from Mendel's second law, the law of independent assortment. The law states that the alleles for two different genes separate into gametes independently of each other.
In plain terms, the way the shape gene sorts has no effect on the way the color gene sorts. When a RrYy parent makes gametes, getting an R does not make Y more or less likely. The two genes are dealt into gametes like two separate coin flips. This independence is what produces all four gamete combinations in equal numbers, and it is why the recombined offspring fall into the 9:3:3:1 pattern.
Independent assortment happens because the two genes sit on different chromosomes, and chromosomes line up randomly during meiosis. Each gene pair is distributed to gametes without regard to the other. Mendel inferred this law from the offspring ratios long before anyone could see chromosomes behave this way, which is part of why his work is so celebrated. The detailed account of how he reached the second law is well documented by Biology LibreTexts.
The practical payoff is this: because each trait assorts on its own, each trait gives its own 3:1 ratio, and the two ratios combine. That insight is the basis of a much faster method than drawing sixteen boxes.
The Faster Way: Probability and the Product Rule
You do not always need to draw a 4×4 grid. The product rule reaches the same 9:3:3:1 answer with simple multiplication, and it scales to more traits where grids become impractical.
Start by treating each trait as its own monohybrid cross. For seed shape, Rr by Rr gives a 3:1 ratio, so any offspring has a 3/4 chance of being round and a 1/4 chance of being wrinkled. For seed color, Yy by Yy also gives 3:1, so a 3/4 chance of yellow and a 1/4 chance of green. Because the traits are independent, you multiply the separate probabilities to get the chance of any combination.
Work through the four combinations:
- Round and yellow: 3/4 × 3/4 = 9/16
- Round and green: 3/4 × 1/4 = 3/16
- Wrinkled and yellow: 1/4 × 3/4 = 3/16
- Wrinkled and green: 1/4 × 1/4 = 1/16
Those four fractions are the 9:3:3:1 ratio, reached without a single box. The product rule is faster, less error prone, and it is the only sane approach once you reach three or more traits. A trihybrid cross would need a 64-box grid, but with probability you just multiply three 3:1 ratios together. This is also how a phenotype probability tool computes results behind the scenes.
The Forked-Line Method
The forked-line method, sometimes called the branch diagram, is a visual version of the product rule. It suits people who prefer to see the combinations branch out rather than multiply fractions in their head.
Draw the first trait as a fork. From a single point, branch into 3 round and 1 wrinkled, the monohybrid ratio for shape. Then, from each of those branches, draw a second fork for color, splitting again into 3 yellow and 1 green. Multiply along each path to the tip. The round branch (3) splits into round yellow (3 × 3 = 9) and round green (3 × 1 = 3). The wrinkled branch (1) splits into wrinkled yellow (1 × 3 = 3) and wrinkled green (1 × 1 = 1). The four tips read 9, 3, 3, 1.
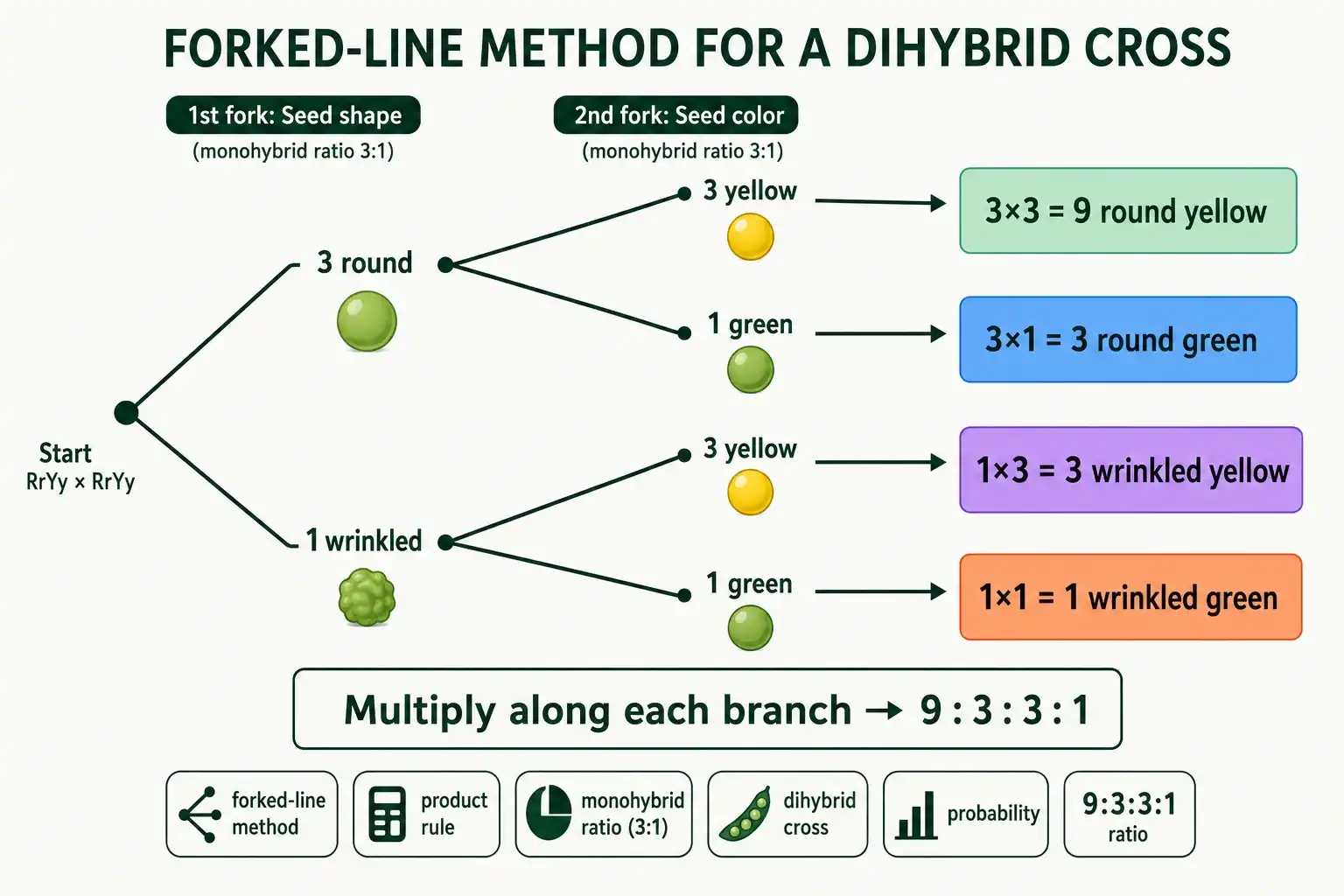
The method shines with three or more traits, where you simply add another fork for each trait and keep multiplying along the branches. It turns a problem that would need dozens of boxes into a tidy tree. For multi-trait crosses, the forked-line method calculator automates the branching and gives you every combination and its probability.
The Dihybrid Genotypic Ratio
The 9:3:3:1 ratio describes phenotypes. The genotype ratio is different and more detailed, because the sixteen boxes contain nine distinct genotypes, not four.
Tally the grid by genotype and you get this breakdown: one RRYY, two RRYy, one RRyy, two RrYY, four RrYy, two Rryy, one rrYY, two rrYy, and one rryy. Written as a ratio that is 1:2:1:2:4:2:1:2:1, and the parts add to sixteen. The double-heterozygote RrYy is the single most common genotype, appearing in four of the sixteen boxes, because there are many independent ways to inherit one of each allele.
This is why naming the ratio you want matters in every dihybrid problem. A question asking for the phenotype ratio wants 9:3:3:1. A question asking for the genotype ratio wants 1:2:1:2:4:2:1:2:1. They come from the same grid but count completely different things, and answering with the wrong one is a common way to lose marks.
Turning the Ratio Into Expected Offspring Numbers
A ratio tells you proportions, but homework and lab work often ask for actual counts. Converting 9:3:3:1 into expected numbers takes one step, and it is the figure you need before testing real data against the prediction.
Add the parts of the ratio first. Nine plus three plus three plus one equals sixteen, so every group is a fraction out of sixteen. To find expected counts, divide your total number of offspring by sixteen, then multiply by each part of the ratio.
Say a cross produces 320 seeds. Divide 320 by 16 to get 20, the value of one ratio unit. Now multiply: the double-dominant class is 9 × 20 = 180, each mixed class is 3 × 20 = 60, and the double-recessive class is 1 × 20 = 20. So you expect 180 round yellow, 60 round green, 60 wrinkled yellow, and 20 wrinkled green. Those four numbers add back to 320, which confirms the arithmetic is correct.
These are expected values, not what you will count exactly. Real offspring scatter around the prediction because inheritance is probabilistic. To judge whether an observed result is close enough to 9:3:3:1 or far enough off to suggest linkage or another interaction, geneticists compare the observed counts against these expected counts with a statistical test. That comparison is where expected numbers become essential rather than academic.
The Dihybrid Test Cross (1:1:1:1)
Not every dihybrid cross gives 9:3:3:1. A test cross, used to uncover an unknown genotype, gives a 1:1:1:1 ratio instead.
Suppose you have a round, yellow plant and you want to know whether it is a true double-heterozygote (RrYy) or carries hidden recessive alleles. Cross it with a fully recessive partner, rryy, which can only contribute the gamete ry. The offspring then depend entirely on which gametes the unknown parent supplies.
If the unknown is RrYy, it makes the four gametes RY, Ry, rY, and ry in equal numbers. Pairing each with ry gives RrYy, Rryy, rrYy, and rryy, which appear as round yellow, round green, wrinkled yellow, and wrinkled green in a 1:1:1:1 ratio. Seeing all four phenotypes in roughly equal numbers confirms the parent was a double-heterozygote. The test cross converts an invisible genotype into a countable result, which is why it remains a standard tool in breeding and genetics teaching.
When the Ratio Breaks: Linkage and Epistasis
The clean 9:3:3:1 ratio depends on assumptions, and real genetics sometimes violates them. When the observed numbers drift far from 9:3:3:1, the deviation itself carries information about the genes involved.
The biggest disruptor is gene linkage. Independent assortment only holds when the two genes sit on different chromosomes. If they lie close together on the same chromosome, they tend to travel into gametes as a unit rather than sorting freely. That biases the gamete ratios and skews the offspring away from 9:3:3:1, with the parental allele combinations appearing more often than expected.
A second disruptor is epistasis, where one gene masks or modifies the effect of another. Epistasis can collapse the four phenotype classes into fewer visible groups, producing modified ratios such as 9:3:4 or 9:7. Both linkage and epistasis are signals worth recognizing, because a distorted ratio is often the first clue that the two genes interact in some way. These topics extend well beyond a basic dihybrid cross, but it helps to know that the 9:3:3:1 ratio is the expectation under independence, not an unbreakable law.
Common Mistakes to Avoid
A handful of errors account for most wrong answers in dihybrid problems, and all of them are avoidable once you know the pattern.
The first is botching the gametes. A RrYy parent makes four gametes, each with one shape allele and one color allele. Writing "RrYy" or "Rr" as a gamete is the single most common mistake, and it ruins everything downstream. Use FOIL and check that each gamete has exactly two letters from two different genes.
The second is making the grid the wrong size. Two traits need a 4×4 grid with sixteen boxes. A 2×2 grid only handles one trait, so if you find yourself with four boxes for a dihybrid cross, you have lost two gametes somewhere.
The third is confusing the two ratios. The phenotype ratio is 9:3:3:1 and the genotype ratio is 1:2:1:2:4:2:1:2:1. State which one you are giving. The fourth is expecting real offspring to match 9:3:3:1 exactly in small numbers. The ratio is a probability across many offspring, and you only see it clearly in large samples, which is why Mendel grew hundreds of plants per cross. A useful summary of these expected ratios and their conditions is maintained by Nature Education's Scitable.
Put the Dihybrid Cross to Work
You now have the full picture of a dihybrid cross. Two heterozygous parents give four gametes each, fill a sixteen-box grid, and produce a 9:3:3:1 phenotype ratio driven by the law of independent assortment. The product rule and the forked-line method reach the same answer faster, the genotype ratio is the more detailed 1:2:1:2:4:2:1:2:1, and a test cross gives 1:1:1:1 to reveal hidden alleles.
Practice a couple of crosses by hand so the gamete logic and the grid become second nature. After that, skip the sixteen boxes and let the Punnett Square Calculator handle any cross from one trait up to five, with every ratio worked out for you. Once two traits feel routine, the same principles scale straight up to trihybrid crosses and beyond.