Mutation-Selection Balance Explained
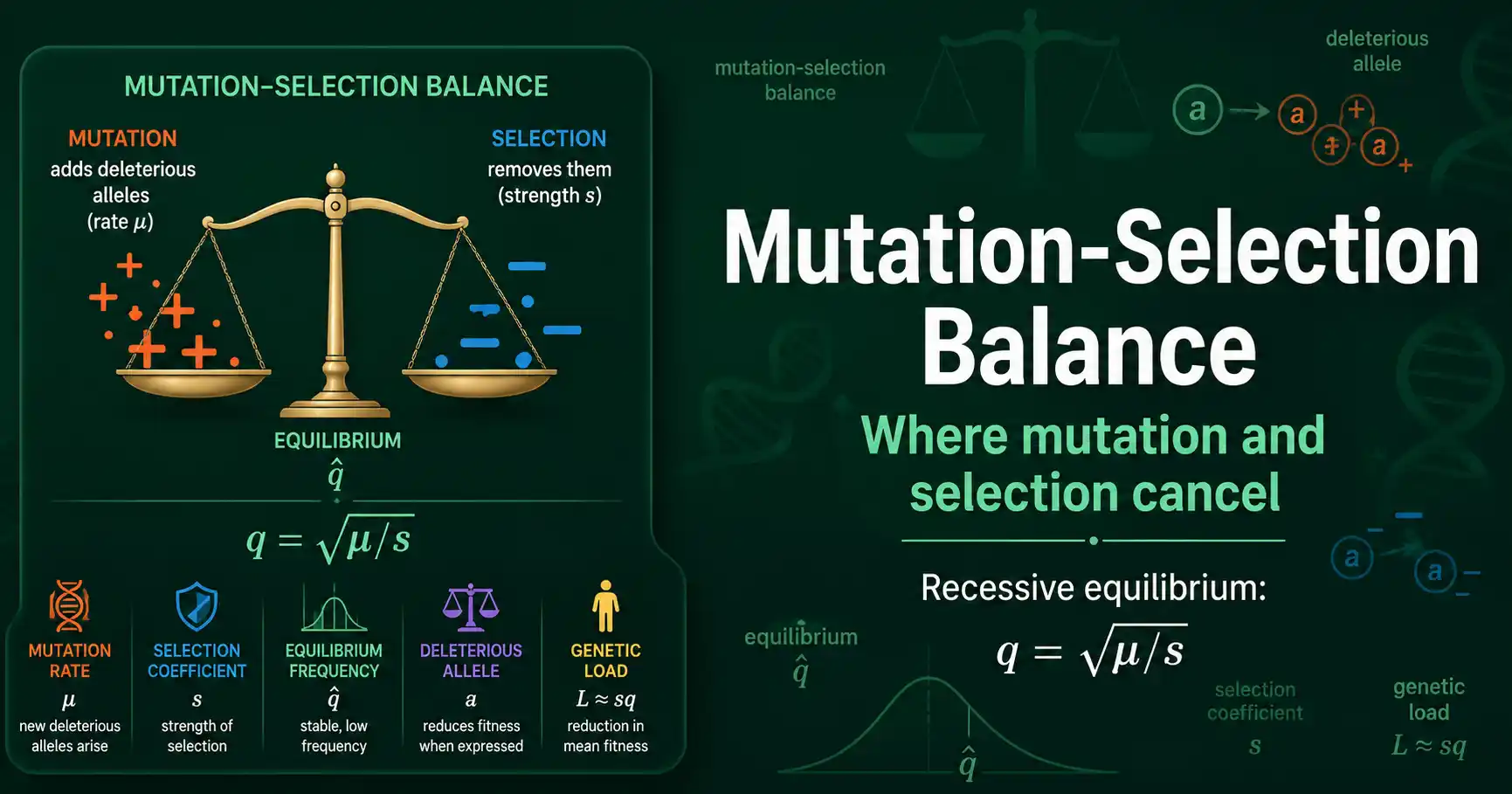
Mutation-selection balance is the equilibrium reached when new deleterious mutations appear just as fast as natural selection removes them. At that point the frequency of a harmful allele stops changing, held at a low, stable level. For a recessive deleterious allele, that equilibrium frequency is the square root of the mutation rate divided by the selection coefficient, q = sqrt(mu/s). This balance is why harmful alleles never quite disappear from a population.
This guide explains the balance and its formulas. It covers how the equilibrium arises, the recessive and dominant cases, a worked example, the concept of genetic load, and why the whole idea matters for genetic disease. It builds on the forces that change allele frequencies in our guide on Hardy-Weinberg equilibrium, with mutation and selection now both switched on.
The Idea: Two Opposing Forces
Mutation-selection balance is a tug of war between two forces acting on a harmful allele. Mutation keeps creating it; selection keeps removing it. The balance is where they cancel.
Mutation is the source. Every generation, new copies of a deleterious allele arise by mutation from the normal allele, at the mutation rate. This steadily adds harmful alleles to the population, no matter how often selection removes them, because mutation is relentless and recurrent.
Selection is the sink. Because the allele is harmful, individuals carrying it leave fewer offspring, so selection steadily removes copies from the population. The strength of this removal is the selection coefficient, s, the fractional fitness reduction the allele causes when its effect is felt.
The equilibrium is where the two rates match. When the number of new deleterious alleles added by mutation each generation exactly equals the number removed by selection, the allele frequency stops changing. That stable frequency is the mutation-selection balance, written with a hat as q-hat to mark it as the equilibrium value. It is low, because selection holds the allele down, but never zero, because mutation keeps replenishing it.
Where the Formula Comes From
The equilibrium formulas are not arbitrary; they fall out of setting the two forces equal. Seeing the derivation makes the square root in the recessive case intuitive rather than mysterious.
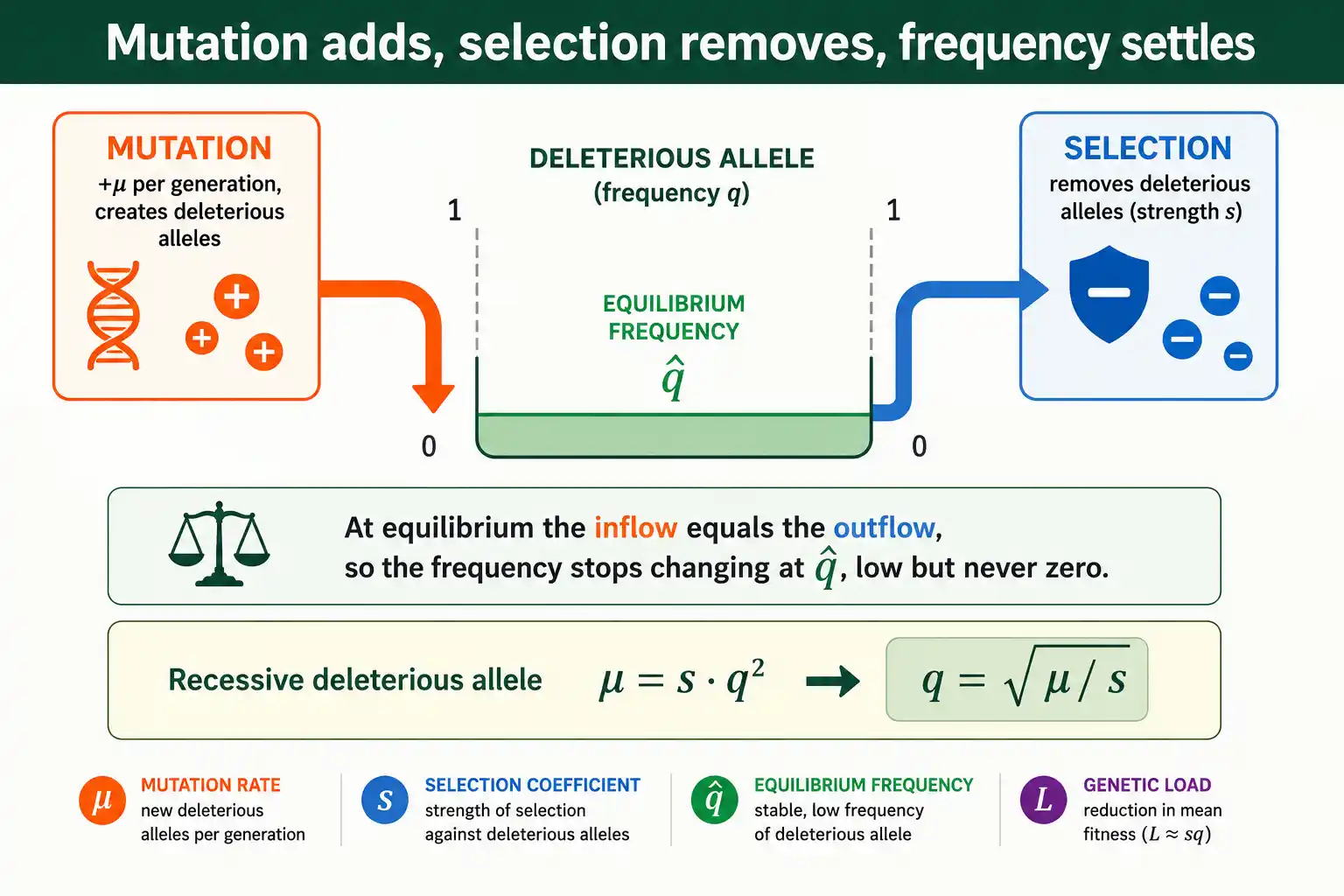
Start with the recessive case. Each generation, mutation converts normal alleles to the deleterious form at rate mu, adding deleterious alleles in proportion to mu times the frequency of normal alleles, which is roughly mu when the deleterious allele is rare. That is the gain from mutation. The loss from selection works on homozygotes, which occur at frequency q squared, each suffering a fitness cost s, so the loss is proportional to s times q squared. At equilibrium the gain equals the loss, so mu equals s times q squared. Solving for q gives q equals the square root of mu over s. The square root appears precisely because selection acts on q squared, the homozygote frequency, not on q directly.
The dominant case follows the same logic with one change. A dominant or partially dominant allele is exposed in heterozygotes, which occur at frequency roughly 2q, and selection removes them with intensity hs. So the loss is proportional to hs times q, not s times q squared. Setting the mutational gain mu equal to this loss, mu equals hs times q, and solving gives q equals mu over hs. No square root, because selection now acts on q directly through the heterozygotes rather than on q squared through the homozygotes. The whole difference between the two formulas comes down to whether selection sees the allele in heterozygotes or only in homozygotes.
The Recessive Case
Most harmful alleles are recessive, so this is the most important case. For a recessive deleterious allele, the equilibrium frequency is the square root of the mutation rate divided by the selection coefficient.
Written out, q-hat equals the square root of mu over s, where mu is the mutation rate and s is the selection coefficient against the homozygote. The square root is the key feature, and it has a clear cause: a recessive allele is only exposed to selection when it is homozygous, which happens at a frequency of q squared under random mating. Because selection only sees the allele in that rare homozygous state, it removes copies inefficiently, so the allele builds up to a higher equilibrium than you might expect.
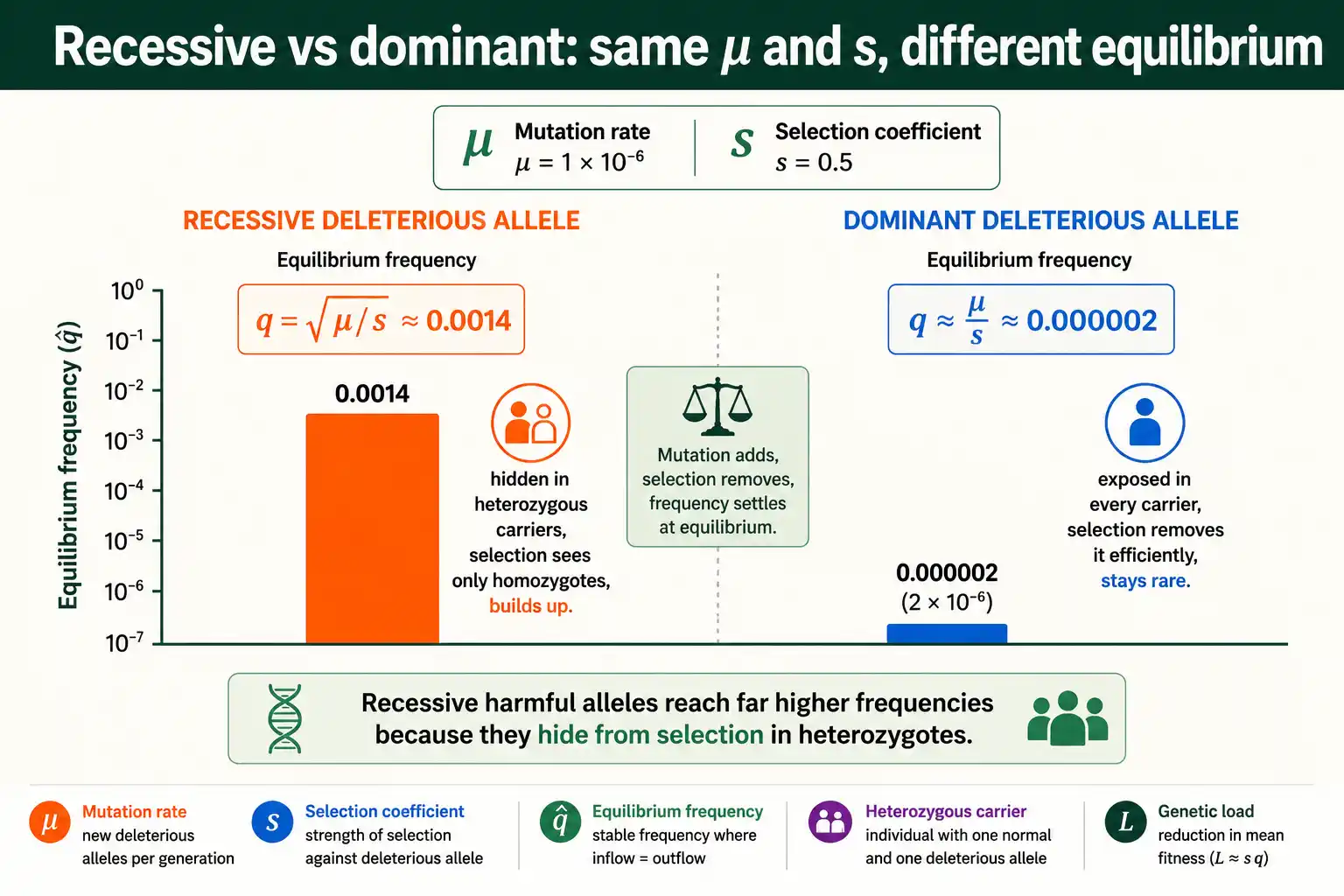
The square root makes recessive deleterious alleles surprisingly common. Suppose a recessive allele has a mutation rate of 1 x 10^-6 and a strong selection coefficient of 0.5. The equilibrium frequency is the square root of 1 x 10^-6 divided by 0.5, which is the square root of 2 x 10^-6, about 0.0014. That is far higher than the mutation rate itself, because the square root of a small number is much larger than the number. Recessive harmful alleles hide in heterozygous carriers, shielded from selection, which is exactly why they persist.
The Dominant and Partial Cases
When the deleterious allele is not fully recessive, selection sees it in heterozygotes too, and the equilibrium is lower. The formula changes accordingly.
For a deleterious allele with dominance, the equilibrium frequency is approximately the mutation rate divided by the product of the dominance coefficient and the selection coefficient: q-hat is about mu over hs, where h measures how much the allele's harm shows in heterozygotes. For a fully dominant deleterious allele, where h is one, this simplifies to q-hat equals mu over s. The square root is gone, because selection now acts on every copy of the allele, not just the rare homozygous ones.
The contrast between the two cases is large. A dominant allele is exposed to selection in every carrier, so it is purged efficiently and sits at the very low frequency mu over s, close to the mutation rate. A recessive allele hides in heterozygotes, so it is purged inefficiently and sits at the much higher frequency sqrt(mu/s). The same mutation rate and selection coefficient produce vastly different equilibrium frequencies depending on dominance, which is why dominance is central to predicting how common a harmful allele will be.
This also explains a real pattern in human genetics. Severe dominant disorders tend to be rare and often arise from new mutations each generation, because selection removes them quickly. Severe recessive disorders can maintain higher carrier frequencies, because the alleles persist hidden in heterozygotes, which connects directly to carrier frequencies and the recessive-disease risk covered in our work on inbreeding and inbreeding depression.
A Worked Example
Put the recessive formula to work on a realistic case. Consider a recessive lethal allele, one that causes death or complete sterility in homozygotes, so the selection coefficient s is 1, with a mutation rate mu of 1 x 10^-6.
Apply the formula. The equilibrium frequency is the square root of mu over s, which is the square root of 1 x 10^-6 over 1, or the square root of 1 x 10^-6. That works out to 1 x 10^-3, or 0.001. So even a recessive allele that is lethal when homozygous settles at a frequency of about one in a thousand, purely from the balance between recurrent mutation and selection.
Now find the carrier frequency, which is what matters for disease risk. Under Hardy-Weinberg, the frequency of heterozygous carriers is about 2q, which is 2 times 0.001, or 0.002, roughly one carrier in 500 people. Meanwhile the frequency of affected homozygotes is q squared, which is 1 x 10^-6, or one in a million. The striking lesson is that carriers vastly outnumber affected individuals, by a factor of about 1 over q, here 1,000 to one. The harmful allele is overwhelmingly hidden in healthy carriers, which is why recessive disease alleles persist. You can explore how the equilibrium frequency shifts with different mutation rates and selection strengths by plugging mutation and selection values into a calculator alongside these formulas.
A Worked Example: Relaxed Selection
The s-in-the-denominator effect is easiest to see with numbers. Take a recessive condition historically held at a selection coefficient of s equals 0.5, with a mutation rate mu of 1 x 10^-6. Its equilibrium frequency is the square root of 1 x 10^-6 over 0.5, the square root of 2 x 10^-6, about 0.0014.
Now suppose treatment makes the condition far milder, cutting the selection coefficient by 90 percent to s equals 0.05. The new equilibrium is the square root of 1 x 10^-6 over 0.05, the square root of 2 x 10^-5, about 0.0045. The allele frequency more than triples, from 0.0014 to 0.0045, simply because selection now removes it less efficiently. The carrier frequency rises in step.
Two things temper this. First, the change is not instant: the allele creeps toward the new, higher equilibrium over many generations, so the effect unfolds across a long time, not within a lifetime or even a few generations. Second, the absolute frequencies remain low, so the practical near-term impact is small. The example illustrates the mathematics of relaxed selection cleanly, the equilibrium scales as one over the square root of s, while showing why it is a slow, long-term consideration rather than an immediate one. It is the kind of effect that matters over evolutionary time, not clinical time.
Genetic Load
Mutation-selection balance produces an unavoidable cost called the genetic load: the reduction in average fitness caused by all the deleterious alleles a population carries at equilibrium. The result here is remarkable and counterintuitive.
The genetic load from mutation-selection balance depends only on the mutation rate, not on how harmful the alleles are. This is the Haldane-Muller principle. For recessive deleterious alleles, the load is approximately equal to the mutation rate u. For alleles expressed in heterozygotes, it is approximately twice the mutation rate, 2u. The selection coefficient does not appear, which seems strange until you see why.
The reason is a cancellation. A more harmful allele, higher s, is removed more efficiently, so it sits at a lower equilibrium frequency, but each copy that is removed costs more fitness. A less harmful allele sits at a higher frequency but costs less per copy. The frequency and the per-copy cost move in opposite directions and exactly cancel, leaving a load that depends only on how fast mutations arise. This principle, worked out by J.B.S. Haldane in a 1937 paper on the effect of variation on fitness and by Hermann Muller, means a population's mutational load is set by its mutation rate, which is one reason the mutation rate matters so much for long-term population health.
The Haldane-Muller principle has a striking practical reading. Because each fully expressed deleterious mutation removed by selection costs, on average, one "genetic death," the total load equals the rate at which deleterious mutations enter the genome. If humans accumulate a meaningful number of new deleterious mutations per genome each generation, the cumulative mutational load is substantial, a point that has driven decades of debate about the deleterious mutation rate in humans, summarized in reviews of the genomic mutation rate and its fitness consequences. The unsettling implication is that the burden does not depend on cleaning up any particular disease; it is set by the genome-wide pace of mutation itself.
The Formulas Side by Side
The two main cases are easiest to compare in a table.
| Case | Equilibrium frequency (q-hat) | Why |
|---|---|---|
| Recessive (h = 0) | sqrt(mu/s) | Selection only sees homozygotes (q squared), so removal is inefficient |
| Partial dominance (0 < h < 1) | about mu/(hs) | Selection acts partly on heterozygotes |
| Full dominance (h = 1) | about mu/s | Selection sees every copy, removal is efficient |
| Haploid | mu/s | Every allele is exposed to selection |
The pattern across the table is about exposure to selection. The more readily an allele's harm is expressed, and therefore seen by selection, the lower its equilibrium frequency. Recessive alleles, hidden in heterozygotes, reach the highest frequencies; dominant and haploid alleles, exposed in every carrier, the lowest. The mutation rate sets the scale in every case, and dominance sets how far above the mutation rate the equilibrium climbs.
Why It Matters
Mutation-selection balance explains why genetic disease never disappears, which is its most important real-world consequence. Harmful alleles are continuously regenerated by mutation, so no amount of selection can fully purge them.
For recessive genetic disorders, the balance predicts the carrier frequencies seen in populations. A recessive disease allele sits at sqrt(mu/s), and most copies hide in healthy heterozygous carriers, which is why recessive conditions can persist at stable frequencies for thousands of generations and why carrier screening finds so many carriers relative to affected people. The balance is the quantitative backbone of recessive-disease genetics.
The principle also has a sobering implication for relaxed selection. If medicine or a changed environment reduces the selection against a harmful allele, lowering s, the equilibrium frequency rises, because the formula puts s in the denominator. Over many generations, weakening selection allows deleterious alleles to accumulate to higher frequencies. This is a real long-term consideration in human genetics, though it plays out far too slowly to matter on any short timescale. Above all, mutation-selection balance ties the mutation rate to the burden of genetic disease, making the rate covered in our guide on what affects the mutation rate a direct determinant of how much harmful variation a population carries.
Frequently Asked Questions
What is the formula for mutation-selection balance?
For a recessive deleterious allele, the equilibrium frequency is q-hat equals the square root of mu over s, where mu is the mutation rate and s is the selection coefficient. For a dominant or partially dominant allele, it is approximately q-hat equals mu over hs, where h is the dominance coefficient, which simplifies to mu over s for a fully dominant allele.
Why don't harmful alleles disappear completely?
Because mutation keeps regenerating them. Even though selection removes deleterious alleles every generation, new copies arise by mutation at a steady rate, so the allele settles at an equilibrium frequency rather than vanishing. This balance between recurrent mutation and selection is why genetic disease alleles persist in populations indefinitely.
Balance, Not Elimination
Mutation-selection balance is the steady state where mutation creates deleterious alleles as fast as selection removes them, fixing their frequency at a low but nonzero level. For recessive alleles the equilibrium is sqrt(mu/s), high enough that carriers vastly outnumber affected individuals; for dominant alleles it is about mu/s, much lower because selection sees every copy. The mutation rate sets the scale, and dominance sets how far the frequency rises above it.
The deeper results are that the genetic load at equilibrium depends only on the mutation rate, the Haldane-Muller principle, and that harmful alleles persist indefinitely because mutation never stops. This makes mutation-selection balance the link between the mutation rate and the burden of genetic disease. To see the applied consequences across evolution and medicine, our guide on mutation rate in evolution and disease carries the story into cancer, viral evolution, and the molecular clock.