Mutation Rate in Evolution and Disease

The mutation rate sits at the center of biology because it sets the pace of change. It supplies the raw variation that evolution acts on, drives the somatic mutations behind cancer, lets viruses outrun our drugs and vaccines, and ticks as the molecular clock that dates the tree of life. The same number, how often DNA changes, runs through all of these.
This guide is the applied capstone of the topic. It covers mutation rate in evolution, in cancer, in viral evolution and drug resistance, and in the molecular clock, showing how one rate connects fields that look unrelated. For the concept itself, our explainer on what mutation rate is sets it up.
Mutation as the Engine of Evolution
Mutation is the ultimate source of all genetic variation, so the mutation rate sets the supply of raw material for evolution. Without mutation, there is nothing for selection or drift to act on, and evolution stops.
The rate matters because it tunes how fast new variation appears. Too low a mutation rate starves a population of the novelty it needs to adapt to changing conditions. Too high a rate floods the genome with harmful changes faster than selection can remove them, dragging fitness down. Most populations sit at an intermediate rate that balances these pressures, an equilibrium explained by the drift-barrier hypothesis in our guide on what affects the mutation rate. To get a feel for how a per-base rate translates into the number of new mutations a genome carries each generation, you can scale a mutation rate up to a whole genome and see how quickly variation accumulates.
This dual role makes mutation a double-edged sword. The same process that generates the beneficial variants behind adaptation also generates the deleterious ones behind genetic disease. Every adaptation in the history of life began as a mutation, and so did every inherited disorder. The mutation rate governs the flow of both, which is why it is one of the most consequential parameters in all of biology.
Mutation Rate and Cancer
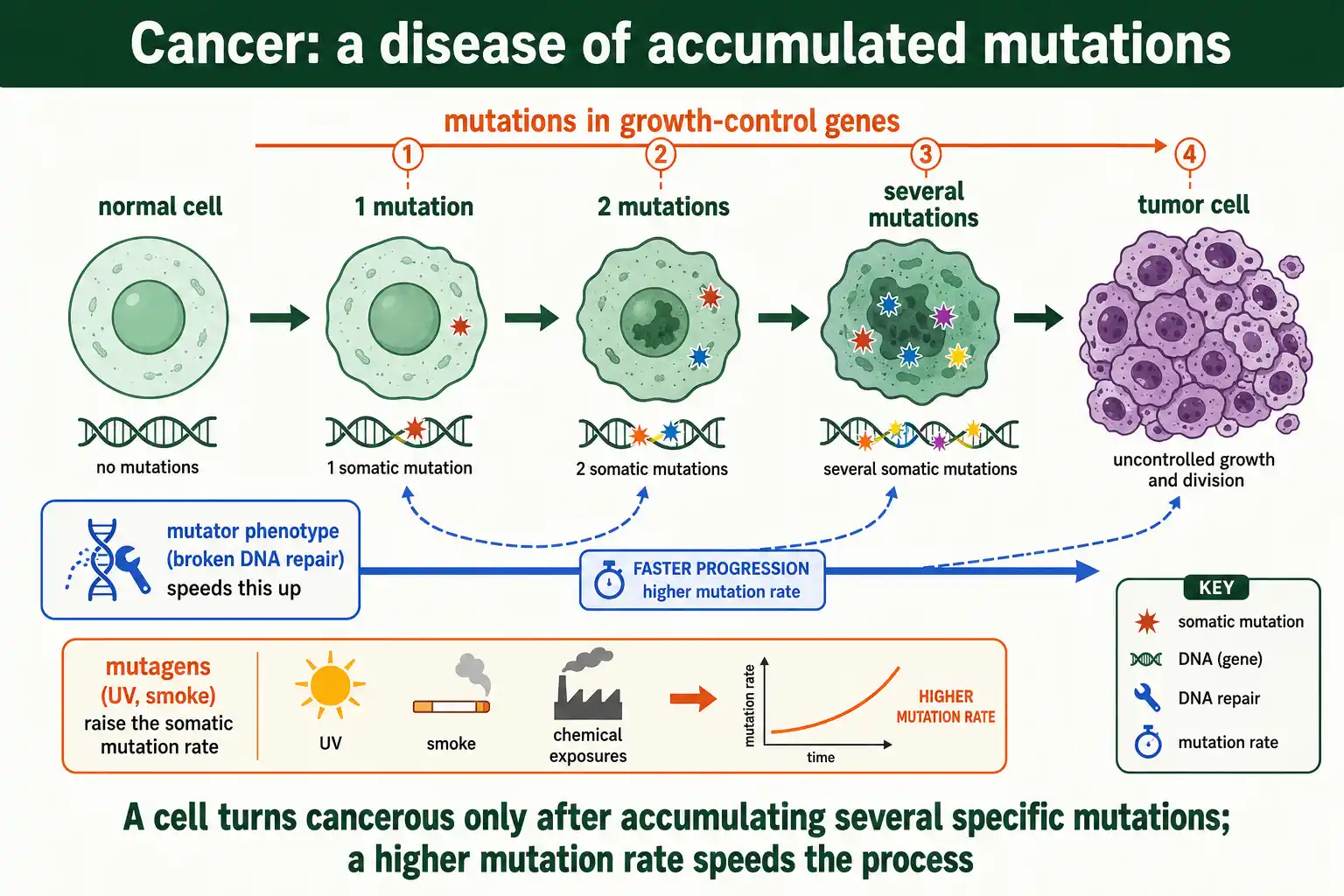
Cancer is, at its core, a disease of somatic mutation, so the somatic mutation rate is central to how cancer starts and progresses. Tumors arise when a lineage of body cells accumulates mutations in the genes that control growth.
The logic is an accumulation problem. A normal cell becomes cancerous only after acquiring several specific mutations, in genes that drive proliferation, disable growth brakes, and evade cell death. Because each mutation is rare, accumulating the full set normally takes a lifetime, which is part of why cancer risk rises steeply with age. Anything that raises the somatic mutation rate speeds this accumulation and raises cancer risk.
This is why mutator phenotypes matter so much in cancer. Many tumors carry defects in DNA repair or proofreading, which send their mutation rate soaring and accelerate the acquisition of further cancer-driving mutations. Inherited defects in repair genes raise cancer risk for the same reason, by elevating the mutation rate across all of a person's cells. Exposure to mutagens, from tobacco smoke to ultraviolet light, works the same way, raising the somatic mutation rate and with it the odds that a cell accumulates a dangerous combination. The connection is direct: more somatic mutations, more chances for cancer. Cancer is a serious medical topic, and anyone concerned about personal or family risk is best served by speaking with a physician or genetic counselor who can assess their specific situation.
There is a twist worth knowing, because the relationship is not entirely one-directional. The same high mutation burden that drives a tumor's growth also makes it more visible to the immune system, because heavily mutated tumors display more abnormal proteins for immune cells to recognize. This is why tumors with very high mutation loads, including those with broken repair systems, often respond better to immunotherapy than tumors with few mutations. The mutation rate, in other words, both fuels cancer and offers a handle for treating it, a duality that runs through this whole topic. Tumor mutational burden has become a real clinical marker used to predict which patients may benefit from certain immunotherapies.
Mutation Rate and Genetic Disease
Beyond cancer, the germline mutation rate sets the baseline incidence of inherited disease, because new mutations are a constant source of disorders. Every generation, the mutation rate guarantees a fresh supply of disease-causing changes.
The clearest case is dominant disorders caused by de novo mutations. Because each person carries dozens of new mutations their parents lacked, some will occasionally strike a gene where a single altered copy causes disease. Many cases of dominant conditions like achondroplasia and some developmental disorders arise this way, in children with no family history, purely from a new mutation in the egg or sperm. The incidence of such conditions is set directly by the mutation rate at the relevant genes, which is why these disorders recur at stable frequencies generation after generation even though selection removes affected individuals. This is mutation-selection balance in action, the equilibrium covered in our guide on mutation-selection balance.
The paternal age effect ties the mutation rate to disease risk in a personal way. Because the male germline keeps dividing, older fathers transmit more de novo mutations, and the risk of certain new-mutation disorders rises modestly with paternal age. The effect is real but gradual, adding a small increment per year rather than a sharp jump. For recessive disorders, the mutation rate works more slowly, replenishing the pool of recessive alleles that selection removes, keeping carrier frequencies stable over long timescales. Across both dominant and recessive conditions, the steady drip of new mutations is why genetic disease is a permanent feature of every population, not something that can be eliminated.
Mutation Rate in Viral Evolution
Viruses, especially RNA viruses, have the highest mutation rates in biology, and that single fact explains much of what makes them dangerous. Their rates can be a million times higher than ours per site.
The numbers are striking. RNA viruses like HIV have mutation rates around 10^-3 to 10^-4 per nucleotide per replication, so vast that a 10,000-base genome accumulates roughly one new mutation every time it is copied. The reason is that the enzymes RNA viruses use to copy their genomes mostly lack proofreading, so errors that a cell would catch go uncorrected, a feature reviewed in work on why RNA virus mutation rates are so high. This high rate is not a flaw from the virus's perspective; it is the source of its adaptability.
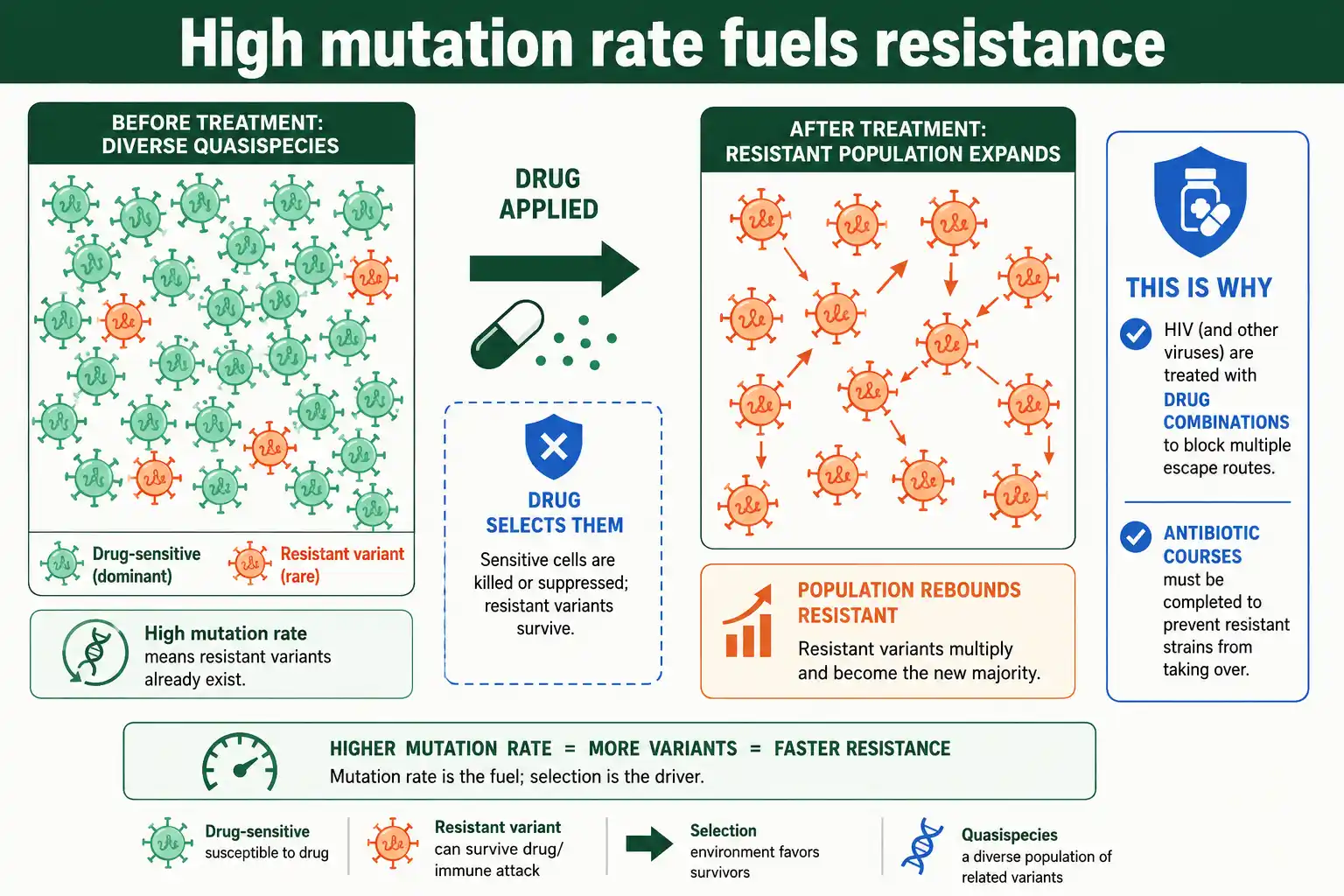
The consequences for medicine are severe. Because a virus like HIV generates enormous genetic diversity, a population within a single patient already contains variants resistant to almost any single drug, which is why HIV is treated with drug combinations rather than one drug at a time. The same diversity lets influenza and other viruses change their surface proteins to escape immunity, which is why flu vaccines must be reformulated and why some viruses evade vaccines. The high mutation rate is the engine behind drug resistance and immune escape alike, the central challenge in treating fast-evolving viruses. This is the same evolutionary process, running at high speed, that shapes the patterns of variation our guide on Hardy-Weinberg equilibrium describes in slower-evolving populations.
Bacteria, Mutators, and Antibiotic Resistance
The same logic that makes viruses evade drugs operates in bacteria, where the mutation rate underlies antibiotic resistance. A higher rate means a faster route to resistance.
In a large bacterial population, even a low mutation rate produces some cells carrying resistance mutations before a drug is ever applied, and the antibiotic then selects them. The bigger the population and the higher the mutation rate, the more likely a resistant variant already exists. This is why resistance can appear quickly during treatment, and why incomplete courses of antibiotics, which leave large surviving populations, are a recipe for selecting resistance.
Bacteria under stress sometimes raise the stakes by becoming mutators. Strains that lose mismatch-repair function have mutation rates up to a hundredfold higher, and such mutator strains are over-represented among clinical isolates of resistant bacteria, because a higher mutation rate speeds the discovery of resistance mutations. The mutator allele then hitchhikes along with the resistance it helped create. This makes the mutation rate itself a target of interest: just as raising a virus's rate can collapse it, ideas for slowing bacterial evolution include interfering with the stress responses that elevate bacterial mutation rates. The mutation rate, once again, is both the engine of the problem and a potential lever against it.
The Error Threshold and Lethal Mutagenesis
The high viral mutation rate has a hidden vulnerability, and exploiting it is a real antiviral strategy. There is an upper limit to how fast a population can mutate and still survive.
That limit is called the error threshold. A virus population exists as a swarm of related genomes, called a quasispecies, clustered around the fittest sequence. As the mutation rate rises, the swarm spreads out in sequence space. Past a critical mutation rate, the error threshold, the population can no longer maintain its genetic information, and it collapses, a phenomenon called error catastrophe. RNA viruses appear to sit just below this threshold, maximizing their adaptability while staying viable.
The error threshold also explains a long-standing puzzle about viral genome size. RNA viruses have small genomes, rarely more than about 30,000 bases, and the high mutation rate is why. A larger genome copied at the same per-base rate would accumulate too many mutations per copy to stay viable, so the high rate effectively caps how large the genome can grow. The viruses that broke this barrier, the large coronaviruses, did so precisely by evolving a proofreading enzyme that lowers their mutation rate, the exception that proves the rule. The mutation rate and the genome size are locked together by the error threshold.
This vulnerability is turned into a treatment called lethal mutagenesis. Drugs that are mutagenic nucleoside analogues raise the viral mutation rate, and pushing it past the error threshold collapses the viral population. A landmark demonstration came from work in Raul Andino's group: Shane Crotty, Craig Cameron, and Raul Andino showed in a 2001 PNAS paper that the drug ribavirin drives poliovirus into error catastrophe by increasing its mutation rate. The principle has since been pursued against influenza, with favipiravir, and other RNA viruses. A mere three-to-fivefold increase in the mutation rate can be enough to tip a virus over the edge, turning the virus's own high mutation rate into the weapon against it.
The Molecular Clock
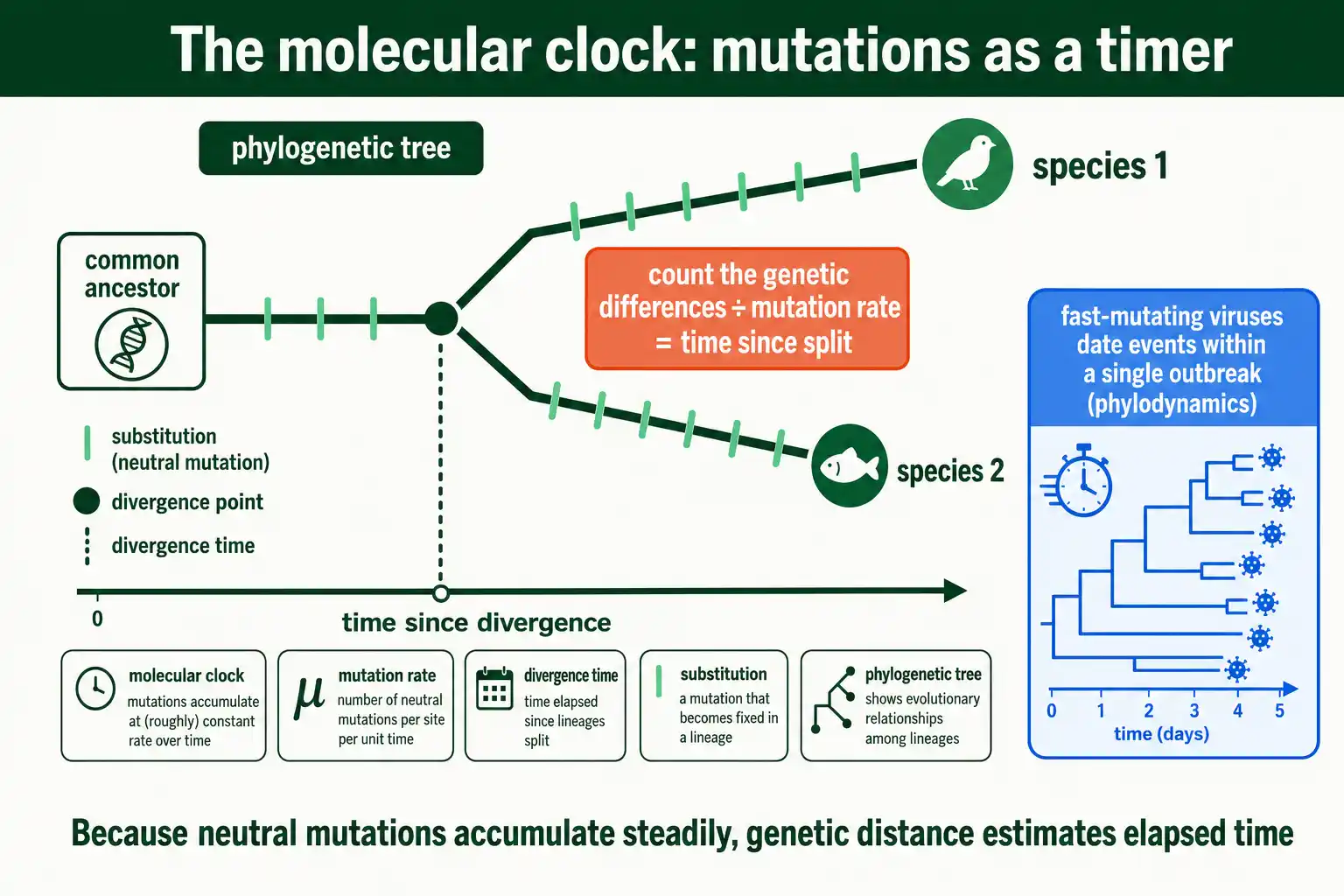
On the longest timescale, the mutation rate acts as a clock that dates evolutionary history. Because mutations accumulate at a roughly steady rate, the genetic distance between species estimates how long ago they diverged.
The logic is direct. If neutral mutations become fixed at a constant rate over time, then counting the genetic differences between two species and dividing by that rate gives the time since they shared a common ancestor. This molecular clock is one of the main tools for dating the tree of life, from the split between humans and chimpanzees to the origins of major animal groups. It works because, for neutral sites, the substitution rate equals the mutation rate, the result explained in our guide on mutation rate versus substitution rate.
The clock is also a practical tool for fast-evolving pathogens. Because viruses mutate quickly, their molecular clocks tick fast enough to date events within a single epidemic, letting researchers estimate when an outbreak began or when a new variant arose from sequence data alone. This field, called phylodynamics, was central to tracking recent viral outbreaks in close to real time. The same mutation rate that makes a virus dangerous also timestamps its spread, turning its genome into a record of its own history.
A concrete example shows the power. By sampling virus genomes over the course of an outbreak and measuring how fast mutations accumulate, researchers can build a dated family tree of the virus, pinpointing when it jumped into humans, how it spread between regions, and when distinct lineages diverged. During recent epidemics, this let public-health teams trace transmission chains and identify new variants within weeks of their emergence, using nothing but viral sequences and the molecular clock. The faster a pathogen mutates, the finer the temporal resolution, so the high mutation rate that makes RNA viruses hard to fight also makes them unusually easy to track through time. It is the same number, read forward to predict escape and backward to reconstruct spread.
One Rate, Many Consequences
The mutation rate connects fields that rarely talk to each other. The table collects its main roles.
| Field | Role of the mutation rate |
|---|---|
| Evolution | Supplies the variation that selection and drift act on; sets the pace of adaptation |
| Genetic disease | Generates the de novo mutations and deleterious alleles behind inherited disorders |
| Cancer | Somatic mutation accumulation drives tumor formation; mutator phenotypes accelerate it |
| Viral evolution | High rates drive drug resistance and immune escape; the error threshold caps them |
| Antiviral therapy | Lethal mutagenesis pushes viral mutation rate past the error threshold to collapse the population |
| Molecular dating | Steady neutral accumulation acts as a clock for divergence times |
The table makes the throughline clear. A single quantity, the rate at which DNA changes, shapes adaptation, disease, infection, and the dating of history. It is generative and destructive at once: the source of every adaptation and every genetic disorder, the engine of both viral escape and the strategy to defeat it. That dual character is why the mutation rate is among the most far-reaching numbers in biology.
Frequently Asked Questions
How does mutation rate affect cancer?
Cancer arises when a cell lineage accumulates several mutations in genes controlling growth, so a higher somatic mutation rate speeds that accumulation and raises cancer risk. Mutator phenotypes, where tumors lose DNA repair or proofreading function, accelerate it further, and mutagens like tobacco smoke and UV light raise the rate and the risk the same way.
Why do RNA viruses mutate so fast?
Because the enzymes RNA viruses use to copy their genomes generally lack proofreading, so copying errors go uncorrected. Mutation rates around 10^-3 to 10^-4 per nucleotide mean a viral genome can pick up roughly one new mutation each time it is copied. This high rate fuels their adaptability, including drug resistance and immune escape.
What is lethal mutagenesis?
Lethal mutagenesis is an antiviral strategy that raises a virus's mutation rate past the error threshold, the point beyond which the population can no longer maintain its genetic information and collapses. Mutagenic drugs like ribavirin push RNA viruses into this error catastrophe. A three-to-fivefold increase in the mutation rate can be enough to drive a viral population extinct.
The Most Far-Reaching Number in Biology
The mutation rate runs through evolution, cancer, viral disease, and the dating of life itself. It supplies the variation that fuels adaptation, generates the somatic mutations that cause cancer, drives the drug resistance and immune escape of fast-evolving viruses, and ticks as the molecular clock that dates divergence across the tree of life. One rate, the frequency at which DNA changes, underlies all of it.
What makes it so consequential is its dual nature. The same process that creates beneficial variation creates harmful variation, so the mutation rate is at once the engine of evolution and a root cause of disease, the source of viral escape and the basis of the lethal-mutagenesis strategy that turns a virus's mutation rate against it. To revisit how that rate is measured, our guide on how to calculate mutation rate covers the foundations behind these applications.